 サイトマップ
サイトマップ研究内容
根の水分屈性発現機構の解明
水分屈性とは、植物の根が水の勾配(水分勾配)に応答して水の多い方向へ屈曲伸長する現象です。植物は自ら動くことの出来ない生物であるため、このように自身の形を生存にとって有利な形にすることで、生存を図っていると考えられます。水分屈性のしくみを理解することで、その性質を利用し、例えば水を有効に利用することの出来る植物の育種や、植物工場などの農業技術の改良など、社会的な要請に応えることが出来ると期待されます。さらに、植物は陸上進出に伴って水を獲得する必要が生じたと考えられるため、水分屈性を研究することにより、植物が地上の環境にどのように適応してきたのか、という進化上の疑問へ応えることが出来ると考えられます。
私達の研究室では、主に植物生理学と分子遺伝学のアプローチから、水分屈性のメカニズムについて研究を進めています。

|

|

|
生理学
水分屈性は、大きく括れば重力屈性や光屈性と同じく屈性と呼ばれる植物の環境応答です。私達は、根はどのようにして水分に応答するのか、また水の方向へ曲がるのはどのようなしくみなのか、そして他の屈性との異同について解析を行っています。
【水の感知】
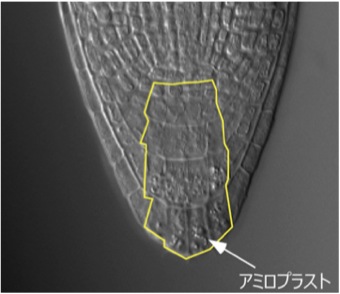
重力屈性においては、根端部のコルメラ細胞(図)内のデンプン粒が沈降することで、重力刺激が感知されています。水分屈性においても、重力屈性と同じく根端部で水の勾配が感知されているのかどうか、外科的な根端の切除や、重粒子線を用いて解析しました。このようにして根端部を除くと、水分屈性を示さない、あるいは屈曲が遅延することがわかりました。この結果から、水分屈性においては根端部に水のセンサーがあると考えられました。今後は、レーザーを用いた1細胞レベルでの研究や、水センサー分子に迫るなど、より詳細な水の感知機構を明らかにして行きたいと考えています。
【オーキシンと水分屈性】
屈性によって根が曲がる時には、植物ホルモンの一つ、オーキシンが根の内部で偏差的な分布が形成されます。根では、オーキシンの濃度が高まると伸長が阻害されるため、偏差的な細胞伸長により屈曲が引き起こされています。このしくみはコロドニー・ウェント仮説と呼ばれ、重力屈性において特に良く調べられています。重力屈性に、PINタンパク質と呼ばれるオーキシントランスポーターが、オーキシンの偏差的な分布形成に必要なことがわかっています。水分屈性では、キュウリの芽生えを用いた実験で、オーキシン応答性遺伝子が、水分の多い側で高く発現していることから、重力屈性と同様なオーキシンを分配するしくみがあると考えられます。一方、シロイヌナズナでは、重力屈性に必須な、PINタンパク質を介したオーキシンの輸送を阻害するNPAやTIBAという薬剤を処理しても、水分屈性は抑制されませんでした。このことから、シロイヌナズナでは、重力屈性とは異なる形でオーキシンが機能していることがわかりました。また、キュウリとシロイヌナズナでそれぞれ水分屈性に必要なオーキシンの役割が異なることから、植物種間で水分屈性を制御するしくみは異なるのではないかと考えています。今後の展開として、まずオーキシンが水分屈性においてどのような役割を持つのか解明すると同時に、様々な植物種における水分屈性の制御について調べていきたいと考えています。
遺伝学
モデル植物シロイヌナズナは、ゲノムサイズが小さく、全ゲノム配列が解読されている上、多数の突然変異体コレクションが蓄積されていることから、分子遺伝学的解析に適した植物種です。水分屈性に関与する遺伝子を明らかにするため、水分屈性を欠損した突然変異体を単離し、その変異原因遺伝子を解析しています。
これまでに2つの遺伝子を水分屈性に必須な遺伝子として同定しています。
【MIZ1】
MIZ1は、機能未知のタンパク質をコードしており、特に根で強い遺伝子発現が見られます。MIZ1を欠損すると、根は完全に水分屈性を示しません。MIZ1は植物に特有のタンパク質であることから、水分屈性が植物の進化に果たした役割を、MIZ1の機能から明らかにしていくことが期待できると考えられます。
【MIZ2】
MIZ2遺伝子は、細胞内において輸送小胞の形成を担うADP-rybosylation factor(ARF)の活性化因子であるグアニンヌクレオチド交換因子(ARF-GEF)の一つ、GNOMをコードしています。このタンパク質は、主に細胞内から細胞膜または細胞外へ分子を輸送するエキソサイトーシスと呼ばれる経路を制御します。特に、オーキシンの輸送を制御するPINタンパク質の局在を制御します。そのため、GNOMを欠損した変異体は、根や子葉の形態に異常をきたしますが、私達が単離したmiz2突然変異体はそのような異常を示しませんでした。そのため、水分屈性に特異的な小胞輸送の経路が存在していることが考えられます。
宇宙
植物は多様な環境刺激の下で生活しています。そのため環境変化に対する応答も、単一の刺激だけではなく、複数の要素を統合した応答になります。屈性においても、重力、光、水分、接触など、多様な刺激によって屈性が起こりますが、植物はそれらの情報を統合し、最終的な屈曲角度が決定すると考えられます。その為、屈性間の相互作用を知ることは、複雑な環境での根の動きを知る上でとても重要です。特に、水分屈性においては、以前から重力屈性との関連について詳しく調べられてきました。
【重力刺激は水分屈性に干渉する】
水分屈性と重力との関連に関する研究は、重力屈性を欠損した突然変異体や、植物に懸る重力方向を撹乱するクリノスタットを用いて行われてきました。このようにして重力の影響を除くと、いずれの場合も水分屈性が強くなりました。つまり、重力は水分屈性を抑制するように働くことがわかったのです。特に、エンドウやキュウリでは、重力屈性の影響を除くことで初めて水分屈性が観察できるため、水分屈性に比べて重力屈性が相対的に強い植物であることがわかりました。
【宇宙実験】
これまで述べたように、重力の影響を除くと水分屈性をより明瞭に観察できることが分かりました。そこで、最も効果的に重力の影響を排除できる環境として、微小重力であるスペースシャトルや宇宙ステーションを使った実験を行っています。現在進行中の実験は、キュウリの水分屈性を観察する実験です。まず一つ目の実験HydroTropiでは、オーキシンと水分屈性との関連に焦点を当てており、根の屈曲に伴うオーキシン応答性遺伝子の分布を解析することで、水分屈性に必要なオーキシンの分配パターンについて、その詳細を明らかにします。もう一つの実験CsPINsでは、オーキシンがどのように分配されるのか、オーキシントランスポーターPINタンパク質の挙動を解析することで明らかにします。このようにして、水分屈性を明瞭に観察できる宇宙実験を行うことで、今後の水分屈性の研究を加速したいと考えています。