文部科学省科学研究費補助金「新学術領域研究」
新学術領域|ゲノム・遺伝子相関







比較ゲノミクス・メチロミクス・トランスクリプトミクスでの
エピゲノム駆動進化の実証
東京大学大学院 新領域創成科学研究科メディカル情報生命専攻 小林 一三
神戸大学大学院医学研究科 内科学講座消化器内科学分野 東 健
基礎生物学研究所 ゲノム情報研究室 内山 郁夫
杏林大学 医学部感染症学 大﨑 敬子
杏林大学 医学部感染症学 神谷 茂
総合研究大学院大学 先導科学研究科・生命共生体進化学専攻 佐々木 顕
基礎生物学研究所 生物機能解析センター 重信 秀治
基礎生物学研究所 生物進化研究部門 長谷部 光泰
千葉大学 大学院医学研究院 福世 真樹
東京大学大学院 新領域創成科学研究科メディカル情報生命専攻 矢野 大和
久留米大学 バイオ統計センター 矢原 耕史
杏林大学 医学部感染症学 米澤 英雄
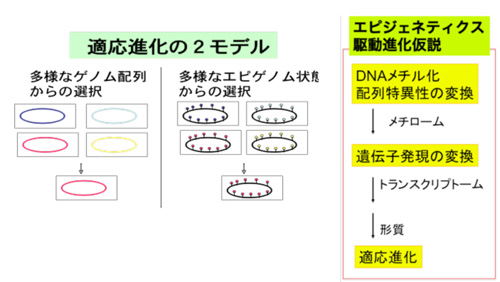
「多様なゲノム配列」ではなく「多様なエピゲノム状態」 を対象に進化が起きるという証拠は、前世紀から植物動物で、そして最近はヒトで集積していますが、その仕組みは謎です。「体細胞イコール生殖系列」と言える単細胞生物である細菌では、この仮説を全ゲノム規模かつ一塩基の分解能で実証することが、今や可能になりました。
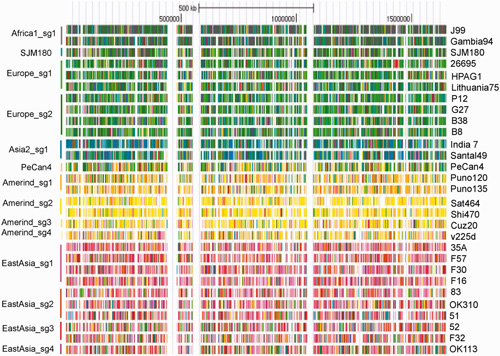
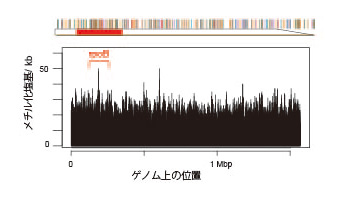
小林班(東大など)は、遺伝と進化のシステムの本質をエピジェネティクスとくにDNAメチル化に求め、細菌をモデルとして、このシステムの両端にあたる「制限修飾系(=配列特異的DNA メチル化酵素+制限酵素)」と「集団ゲノム」を解析し、「DNA 配列認識ドメインの遺伝子内の移動と種を超えた移動」「遺伝情報伝達の痕跡からの種内集団構造の推定(染色体ペインティング、図)」を達成しました。これらから、「制限修飾系が、有無と認識配列と発現を変換することによって、多様なエピゲノム(メチローム)を実現し」、それらは「多様な遺伝子発現パターンと形質セット」に対応するとともに、「遺伝情報の流れからの隔離」をも規定し、「自然選択の単位となって、適応進化が実現する」という「エピゲノム駆動進化仮説」 を提唱しました(図)。全ゲノムのメチル化状態を、PacBio シーケンサーで一塩基の分解能で解明し(図)、この仮説の検証に近づきました。
この「エピゲノム駆動進化仮説」を、近縁オーミクス比較によって実証することが、本研究の目的です。胃がんを起こすピロリ菌を中心に、近縁系列間で、ゲノム・メチローム・トランスクリプトームを比較し、進化過程を追跡し、適応との関連を探ります。さらに、これを制限修飾系の活動の内外環境への応答と結びつけ、適応進化が選択(ダーウィン進化)だけでなく、環境への能動的な応答(ラマルク進化)による可能性をも検討します。
本研究の成果は、遺伝と進化を巡る2世紀の宿題に答え、生命の理解に大きなインパクトを持つでしょう。また、それはゲノム配列でなくエピゲノム状態を標的とする「エピゲノム育種」への道を開くでしょう。
![]()