文部科学省科学研究費補助金「新学術領域研究」
新学術領域|ゲノム・遺伝子相関







月別アーカイブ
計画研究班別アーカイブ
公募研究班別アーカイブ
旧公募研究班別アーカイブ
「研究経過報告」内を検索
【名古屋大】金岡班の記事を表示しています
リンク:研究室に行ってみた チューリッヒ大学 進化生態ゲノミクス 清水健太郎 文・写真・川端裕人
ノンフィクション・フィクション作家である川端裕人さんによるNational
Geographic日本版(Webナショジオ)の連載「研究室に行ってみた」で研究を紹介していただきました。2015年3月7日から17日までに7回連載予定で、
第1回 「新種誕生」を見にスイスアルプスへ行ってみた
第2回 わずか100年でどのように新種が誕生したのか
につづき、次世代シークエンサー、シロイヌナズナ属異質倍数体ミヤマハタザオのゲノム解析、自殖の進化、雄の進化などがトピックです。
![]()
細胞工学の連載「ゲノムで進化の謎を解く!」に寄稿しました。
第8回 20世紀以降にも種分化は起きている:ゲノム重複による新たな環境への適応
過去150年にスイスの小さな村で現れた倍数体新種形成の顛末と、倍数体ゲノム・トランスクリプトーム解析用ソフトウェアHomeoRoq、シロイヌナズナ属異質倍数体ミヤマハタザオのゲノム解析などについて解説しました。
文献情報
![]()
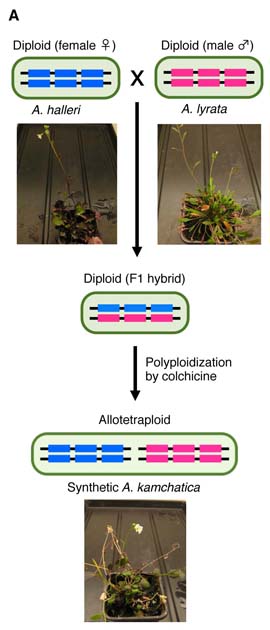
近縁種の2つのゲノムが1つの細胞内に共存する状況は、雑種形成、受精後生殖隔離、そして異質倍数体種分化でのゲノム相関を理解するための鍵である。しかし、これまで近縁種間でのゲノム配列の高い相同性のために、配列を区別して解析することが困難であった。我々は東工大情報系の瀬々研究室との共同研究により、次世代シークエンサーを用いて雑種・異質倍数体の遺伝子配列・ゲノムを解析するためのバイオインフォーマティックワークフローHomeoRoqを開発した。
とくに、2つの異なる種が交雑し,かつ倍数化を起こした異質倍数体種の解析にこの手法は有用である.通常の種間雑種は親の染色体を1本ずつ(1セット)持っているが,異質倍数体は四倍体では2本ずつ(2セット),もしくはそれ以上を持つのが特徴である.このように染色体のセット数が増えるという現象は,生物全般に普遍的に見られるもので,特に植物では頻繁に起こることが知られる.現存する全ての被子植物種の歴史上にこのような倍数化が複数回起こり,それが植物の遺伝的・形態的多様性を高めるのに役立ったというのは,これまでの進化ゲノム学的な知見でも明らかにされている.
比較的最近に起きた異質倍数化では,その両親種と異質倍数体種の両方が現存することが多い.場合によっては両親種の好む環境が明確に異なっていることも多い.このような場合に,これらの種の好む生育環境または地図上の分布域を比較してみると,異質倍数体種は,両親種の中間的な環境,または両者を包合する分布,つまり親種よりも広い分布をとる傾向がある.ミヤマハタザオArabidopsis kamchaticaは、比較的高温の環境に生育するハクサンハタザオA. halleriと低温環境に生息するのセイヨウミヤマハタザオA. lyrataを両親とする異質倍数体であり、広い緯度の海岸から高山まで多様な環境に生育している。異質倍数体はどのようにして,そのような親種とは異なる環境への耐性,またはより広い分布を手に入れたのだろうか?
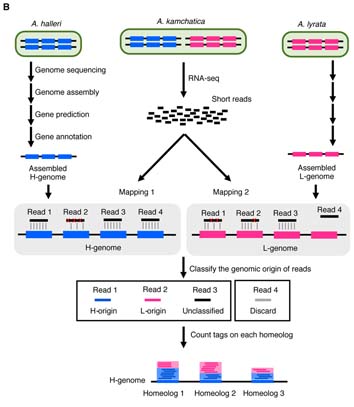
この問いに答えるためには,異質倍数体のゲノムワイドな遺伝子発現解析が必須であった.異質倍数体は同じ1つの遺伝子を2種類ずつ(それぞれの親から一種類ずつ)受け継いでいる.これらの由来親の異なる2種類の遺伝子は,ホメオログ遺伝子と呼ばれる.つまり,異質倍数体は両親種よりも発現させる遺伝子の数,または遺伝子の組み合わせの選択肢が多い.このように,遺伝子の発現制御が異質倍数化によって変化し,それが両親種とは異なる環境耐性を手に入れる鍵となった可能性が高いと考えられる.しかし,どちらのホメオログ遺伝子がどれだけ発現しているのかを知るためには,従来の解析方法では不十分であった.ホメオログ遺伝子間の違いはごくわずかなので,そのわずかな違いを手がかりに由来親を特定する方法が確立していなかったためである.
まず我々は,親種のゲノムアセンブリを行い,それぞれのホメオログの配列情報を得ることから開始した.また,両者のクオリティの違いによるカウントバイアスを避けるために,同じRNA-seqリードを別々にそれぞれのゲノムにマッピングして片方にしかマップされない物は除外し,ミスマッチ数の少ない方を正しい由来親と認定するという方式をとるなどの工夫を凝らした.その結果,人工的に作成した異質四倍体Arabidopsis kamchaticaのRNA-Seqでは,88%以上という高い確率でリードの由来親を特定できるようになった.さらに、PyroMarkを用いてホメオログの発現比率を実験的に測定したところ、HomeoRoqの結果とよく一致した。また,従来の論文では,限られた遺伝子数の発現比較から,異質倍数化後にはストカスティックに大規模な遺伝子発現変化が起こると報告されていたが,ラボ環境で育てたこの人工四倍体では,従来いわれていたほど大規模な不規則な変化は起こっていないことが確認された.
さらに、統計的な過分散をとりいれて、環境変化によってホメオログ間の比率が有意に変化した遺伝子を探索する手法を開発した。人工倍数体を低温ストレスにさらしたところ、ゲノムの約1%の遺伝子でホメオログ間の比率が処理前後で有意に変化しており、RD29B, COR15などシロイヌナズナでよく研究されている低温マーカー遺伝子が含まれていた。この結果は、異質倍数体が、それぞれの親に固有のストレス応答シス配列を受け継ぎ、広い環境耐性を得たことを示唆する。
このパイプラインは,今後,動物・植物・菌類の雑種・異質倍数体の発現解析のみならず、倍数体ゲノム配列解析に適用可能である。さらに、実際の野外の植物個体がどのように親のゲノムを使い分けて環境適応しているのかを調べるための有力なツールとなる.
Genome-wide quantification of homeolog expression ratio revealed nonstochastic gene regulation in synthetic allopolyploid Arabidopsis
Satoru Akama*, Rie Shimizu-Inatsugi*, Kentaro K. Shimizu**, and Jun Sese** (* equally contributed, ** corresponding authors)
Nucleic Acids Research 42: e46 (2014), doi:10.1093/nar/gkt1376
![]()
東南アジア熱帯雨林では、フタバガキ科などの数百種もの樹木が、数年に一度、不規則な間隔で一斉に同調して開花します(写真)。しかしながら、なぜこのような大規模な開花が誘導されるのか、その仕組みについては長らく謎とされてきました。そのため一斉開花は、熱帯生物学で最も壮大でミステリアスな現象と言われ、世界中の研究者の注目を集めてきました。開花誘導の仕組みについては、乾燥、低温、日照といった環境要因に加え、栄養蓄積量などの内部要因が開花を引き起こすとの説がこれまでに提唱され、長く論争が続いてきました。今回我々は、2009年にボルネオ島ランビル国立公園で起きた一斉開花の際に、フタバガキ科樹木Shorea beccariana(写真)の網羅的発現解析を行い、遺伝子発現の観点からこれらの仮説の検証を行いました。
50mにもなる高木からのサンプル採集は、国立環境研究所の竹内やよい博士、筑波大学の田中健太博士らとの共同研究により、日本の研究グループによって設置された高さ80mを越えるクレーン(写真)を用いることで可能になりました。非モデル生物であるShorea beccarianaの発現解析にはRoche 454 次世代シークエンサーを用い、一斉開花をはさむ4時点のサンプル間で1128遺伝子の発現が変わっていることを確認しました。花芽形成に関わる遺伝子に注目すると、その翻訳産物が花芽形成を誘導する植物ホルモン「フロリゲン」であるFT (FLOWERING LOCUS T)遺伝子のホモログの発現が、一斉開花の約3週間前の雨量が非常に少なくなる時期を境に上昇していることがわかりました。またFT遺伝子を直接抑制し、花芽形成を抑えるMADSボックス遺伝子SVP (SHORT VEGETATIVE PHASE)のホモログの発現が同様に雨量の少なくなる時期を境に低下していました。これらの遺伝子をシロイヌナズナで強制発現したところ、それぞれ花芽形成時期を促進させる、あるいは遅延させるといった表現型を引き起こしたことから、花芽形成に関わる機能を保存しているものと考えられます。さらに、1128遺伝子をシロイヌナズナの発現データベースと比較したところ、長期の乾燥で発現変化する遺伝子の多くが、この雨量の少なくなる時期に発現変化していることがわかりました。これらの結果は、植物が実際に乾燥を感じ、その後、花芽形成に関わる遺伝子の発現が変化を始めることを示唆しており、「乾燥が一斉開花を誘導する」という仮説を支持します。東南アジア熱帯雨林では、地球環境変動の結果、より頻繁に雨量の少ない時期が生じることが予想されています。このことは、一斉開花の規模、頻度を変化させる可能性があり、今後注意深い観察が必要になってくると考えられます。これまで、一斉開花時期の予測の難しさから、たくさんの種子を計画的に得ることができず、このことが熱帯雨林再生の一つの障害となっていました。しかしながら、本研究を応用し、発現解析によって開花、種子形成時期の予測ができるようになると、これらの保全の問題の解決にも貢献できる可能性があると期待しています。
Kobayashi, M.J., Takeuchi, Y., Kenta, T., Kume, T., Diway, B., Shimizu, K.K. (2013) Mass flowering of the tropical tree Shorea beccariana was preceded by expression changes in flowering and drought-responsive genes.
Molecular Ecology, Early View
http://onlinelibrary.wiley.com/doi/10.1111/mec.12344/full



(班友 チューリッヒ大学 清水健太郎)
![]()
![]()
![]()
文部科学省科学研究費補助金「新学術領域研究」